One of the coming developments I’ve been the most excited about in recent years are Groupnets (what I originally called “symbiont networks” in a 2004 article). Groupnets are tightly connected, sensor-equipped groups of individuals that automatically share rich and nearly 24/7 continous data (pictures, audio, video, data) about their lives to each other, allowing them to watch each other in realtime, review and relive recent events, and even speak words of encouragement into each other’s ears as appropriate. The group will be there for individuals when in they’re in need, and these folks will truly begin to see the world through each other’s eyes.
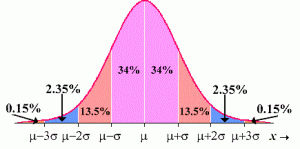
Because there are usually only a few people who are out on the far edges of the normal curve of emotions, thought, and behavior at any time, and a far larger number of others closer to the center of human feeling, thought, and behavior, this kind of tight connectivity always tends to keep folks out on the edge from engaging in too much extreme, dysfunctional, or destructive behavior, gently pulling them back toward the center of the group over time. When an extreme person talks in an emotionally open way to a normal person, they each engage their mirror neurons and related brain systems to internally represent other people’s feelings and thinking within their own heads. The more intimate and frequent social interaction becomes, the more the most adaptive ways of thinking (usually in the center, but occasionally on the edges of human thought and behavior) takes over, in the minds of any member of a cognitively diverse group. To increase the power of therapeutic social interactions, particularly dysfunctional people may need to be attached to particularly large but still intimate groups (for more on these claims, see the books below).
Over the next few decades, I’m convinced that Groupnets and the AI-enabled platforms they occur within will become the most powerful group-based emotional cognitive behavioral therapy we can use on a daily basis to continually improve our lives. They’ll be tremendously empowering for professional and self-improvement, for family and mental health therapy, addiction therapy, rehabilitation in prisons and during parole, any area where feeling, thought, and behavior change is particuarly difficult. Below is a brief 2008 video I did on the idea.
All the shared data of our groupnets will get autoarchived inside private social networks of highly emotionally and values-connected individuals, and autosummarized, like the way the AI behind Google Photos will now automatically find the most interesting moments in your uploaded video for sharing with others. These social networks will be our “Private Facebooks”, the Private Social Networks that many of us are still waiting to emerge, built to encourage sharing and strengthen the power of social bonds among our most intimate friends and family. That will be a social network that has all the core sharing capabilities we need, whose interface we can extensively control and customize with evidence-based insights from social psychology, and whose owners aren’t focused on advertisers over users.
Just as with desktop computers, whose incremental new features no longer held much interest for us once they reached a certain level of functionality in the mid 2000’s, allowing us to turn away from them toward more intimate and portable laptops, then tablets, then our phones, I think we’ll soon tire of additional “features” in the Facebook of today. When we want to do deep personal sharing on social platforms, I expect many of us will turn increasingly away from ad-driven corporate media social networks to Private Social Networks and Groupnets built by companies that share our particular values. Once these deep social sharing capabilities get commoditized, just as with desktop computers, Facebook, at least the kind of Facebook that exists today, won’t be the coolest thing on the block anymore. It will increasingly be seen as just another kind of ad-driven media property: something lowest common denominator and commercialized that we dip into only on occasion, and increasingly leave behind as we grow up.
What we really want to connect better on this precious planet right now are people, even more than gadgets. Think the Internet of Things is cool? Subscribe to @internetofshit for some hilarious tweets on how naively we think about all our silly gadgets today, and how much they make the marketers salivate. An Internet of Families and Close Friends will be way more helpful and meaningful to most of us today, in these final decades of weak AI, than an Internet of Things, though of course we want to keep growing both as fast as we can. The coming Internet of Families will be all the home, mobile, and wearable sensors and devices we’ll use to enrich and make realtime our intimate Private Social Networks, and increasingly turn them into Groupnets.
Being more intimately connected to key individuals, both family and work, in ways that are unobtrusive and asynchronous, like texting, with AI filtering and autosummarizing everyone’s data streams, and which allow us to make “bids” for synchronous communication with various people, at special places and times in the day that they or their groupnet have set aside to be interrupted, will allow us both to focus better when we work, turning off all our feeds except for emergency messages, and when we need a break and crave some social connection or ability to help, to immediately be more intimately connected to those that matters most to us.

Side Note: Read Jeff Sutherland’s excellent Scrum, 2014, to understand that an ideal way to get more and better things done in less time is to organize our days, and our weeks, into natural rhythms of uninterrupted sprints, working with small (9 or less) crossdisciplinary teams, punctuated by short, and social, recoveries, “hangouts” in physical and virtual space whose times and places we’d love to leave to software that organizes them for us. The best of the coming groupnets will use AI to help us find and protect our natural work/social cycles, while greatly increasing the intimacy and the power of our networks.
For each of us, there are something much less than one hundred and twenty special people that we want to have far better archives on, and better tools for helping each other communicate, grow, and thrive. I’ve argued previously that a Groupnet with a Dunbar number (120 or less) of individuals that are deeply emotionally connected via digital systems, share common core values, yet are also both cognitively and skills diverse, will soon greatly outperform unconnected individuals (“solitaries”). All that’s needed are more people-centric platforms and more reliable and affordable rich digital connectivity. Both of those ingredients should be here soon.

For more on why it is that cognitively diverse groups outcompete cognitively narrow ones when working on any of life’s “hard problems” (poorly structured, ambiguous, complex, competitive), see Scott Page’s, The Difference: How the Power of Diversity Creates Better Groups, Firms, Schools, and Societies, 2008, and Markova and McArthur’s Collaborative Intelligence: Thinking With People Who Think Differently, 2015. Both of these are excellent introductions to the future of collaboration. For an example of what life looks like when you don’t have cognitive diversity in your group, read Nick Carr’s The Shallows, 2011, or Eli Parisiner’s The Filter Bubble, 2012. If you don’t set your filters to include people and ideas that are valued by others who think differently from you, you’ve got a big hole in your world simulation, and others will cheerfully keep exploiting that hole to take away your cake, until you finally wise up and improve your world view.
I’ve been excited to see live streaming apps like Periscope and Meerkat take off in recent months. These are a huge step toward Groupnets. Thanks Twitter for recognizing the incredible new intimacy, empathy, support, and emotional connectedness that live streaming offers. Let’s hope Google Fiber (wired), Project Fi (wireless), and other consumer-oriented initiatives continue to make progress against America’s anticompetitive telco and media oligopolies, which are building out their networks as slowly as they can, trying to delay the inevitable rise of peer to peer media, and all the empowering (and less financially controllable) new Little Guy business models that will allow.

Remember Path? It is a Private Social Network that launched in 2010. Path allows up to 150 users to do intimate photo and message sharing with close friends and family. Its functionality has always been very limited (they didn’t do too much development with that $75M in VC money, apparently). Nevertheless it’s user base has grown, particularly outside the US (for example, 4 million Indonesian users by 2014) perhaps in places where customers were less picky about what was available. In 2014 Path got acquired by a South Korean internet company, Daum Kakao. South Korea’s bandwidth is two to three times better than ours, so with luck we may see Path do some interesting things as a Private Social Network in the next few years. Or not. We shall see.
South Korea, Japan, and a handful of other countries now rolling out 5G networks are places where you could build a first gen Groupnet platform today. They have better government guidance, less corporate lobbying slowing down the emergence of real internet competition, greater population densities, and other factors that combine to keep them beating the US at the critical public good of a dependable, cheap, blazing fast internet, the backbone for our digital life. Launch your Groupnet apps there first, and you could achieve First Mover Advantage.

Tinybeans is another deep sharing Private Social Network startup. It started in 2014, and doesn’t have very much funding yet ($1.75M on Crunchbase). But it’s management is taking a very clever approach. They are focusing the platform on sharing of children’s photos, among families. Targeting children’s photos, videos, and accomplishments seems a smart way to build a passionate Early Adopter core for a deep Private Social Network. Most of us are particularly sensitive about sharing children’s photos, so we don’t share many on Facebook, which has low user control and trust. As long as Tinybeans can navigate the legal issues with storing data on underage users, provide good functionality and prioritize security, privacy and control, they should grow nicely.
There are other examples of rich sharing private networks, like SmugMug for professional photography. But I don’t know of any other Private Social Network platforms that are designed for deep sharing among small groups, with features that promote social bonding, intimacy, group transparency, and human development. Please let me know if you do.

NextDoor has the potential to become this kind of social network, for a subset of their members. At first, I could see them helping folks to put more webcams on their front lawns, making our neighborhoods more transparent and secure — an easy sell to security-minded folks. Next they could help an internet of families, with its sensor packages and online archives, to emerge between those neighborhood homes that want greater intimacy between the children and adults in their various families. That’s a much more exciting and socializing development. But to do that, they’d have to make it easier for families that want to make friends with other local families to build out deeper profiles, and more easily find each other by interests, skills, projects, heritage, religion, political beliefs, age of children, and other search options, and set up easy ways for people to interact (dinners, hangouts) with each other in friendbuilding activities on their network. [2023 Note: I doubt NextDoor execs ever had an interest in any of this. It’s messy, exposes them to lawsuits, and has no relation to their extractive, soulless ad-driven business model.]
I think if Facebook was serious about improving social interaction, they’d buy or build something like NextDoor. They’d also buy or build something like Neighborgoods, a platform that allows local families to list things they’re willing to share, a good way to get people out of the solitary, consumption economy. Google could build out these local family tools in G+, and gain a few more points of market share growth (5% of over the next 5 years?) against Facebook. But will they? Even LinkedIn could build something powerful and personal to support local small businesses.
Big social network platforms seem lukewarm about building out stronger local communities. They also aren’t really in the business of helping people to connect more intimately, or supporting their psychological development. Perhaps this is because such efforts aren’t easily monetizable beyond goodwill, and can get messy, political, and controversial. So it will probably fall to the entrepreneurs to keep moving us toward this future.
But I think any good survey would show a substantial fraction of people want much better and richer connections with family and friends. Someone’s going to take seriously this deep common desire for greater intimacy, and build us an Internet of Family and a rich-featured deep sharing Private Social Network, starting with some Early Adopter families and their closest friends. It will grow rapidly from there. While writing this post I searched “Internet of Family” and see that others have come to the same conclusion. For example, see “The Internet of Family and Friends Will Rule the Future“, CMO, 7.21.15, by Tim Joyce of The Cambridge Group. These amazing advances in human connectedness and empowerment are coming. We want them, so it’s just a matter of time. I hope you’ll let us know in the comments wherever you see deep sharing Private Social Networks and an Internet of Family emerging, and what we can do to make them both come faster. One last thing.
Our Global Family, Biological and Adopted

Once the Internet of Family and rich Private Social Networks exist, it’s easy to see we’ll not only be able to stay deeply connected to our own biological family and friends around the globe, we’ll be able to easily reach out to families that we don’t know. It’s exciting to realize that among the those tens of millions of priviliged folk who have a strong desire for social justice, a good deal of that digital reaching out will be to the most at-risk families who really need our help. These new networks will help us to find and help others in very personal, one-to-one ways. By befriending and supporting a small group of others through them, we’ll also greatly help ourselves grow in personal effectiveness and wisdom.
I think Family Adoption, being encouraged to adopt (commit to long-term support) a family that is in great need, not just for an isolated encounter over the holidays, as happens commonly today, but for the years of childraising of the children involved, will become a new phenomenon on these platforms. Family adoption discovery systems will be built into all the better Private Social Networks and Groupnets, and the Internet of Family will make that adoption feel like we are bringing those individuals into our own homes. The death of distance that comes from affordable broadband and wireless and great sensors will give us the ability to really help, one on one, those folks anywhere in the world who need our help when they need it the most, the way we try to do within our own families today.
Handup.org, whose CEO and co-founder is my friend Rose Broome, runs a fantastic new charitable giving platform that offers direct ways to improve the lives of our homeless neighbors and other low-income local residents. Individuals are verified as needy by various community and homeless services groups around the country. At their Help Someone tab, you have the ability to directly help individual local people in need, and search their database by first name, neighborhood, or basic needs. 100% of your donation goes to your selected members needs. This is a fantastic way to give directly to those in need. In coming years, I look forward to seeing more intimate and realtime ways of connecting to any of the individuals in this database who might want such contact from their community of supportive “fans.”
Now imagine a service like Handup, launched today by a social entrepreneur in the slums next to any major emerging nations metropolitan area, like Lagos, Nigeria or Rio de Janiero, Brazil. [2023 Note: Or imagine such a service launched in all of the refugee camps around the world. UNICEF says 17 million children under 18 are presently growing up in such camps, with very limited living conditions, prospects and mobility. The World Bank says the average time spent in refugee camps is 10.3 years. The number of displaced children, forced from their homes by various events (war, famine, pandemic, climate, etc.) reached a new high of 43 million. These are large absolute numbers, but still very small relative to the billions of us who are not displaced, and are able to help.]
Such a service would offer us a searchable adoption needs database with thousands of families in need, and encourage each of us to adopt at least one of these families for a minimum period of time (five years?), and ideally stay intimately connected to them for a lifetime, or at least for the entire childhood of the children (theirs and ours), in a Private Social Network with an Internet of Family connecting us. Services to help our children learn each other’s languages and culture would be part of the platform, as well as a host of contract services to help the families in need. Visitors to the site would be encouraged to donate to help volunteers build the adoption needs database in countries and cities not yet represented. How many families do you think this service might help? Are there any such platforms today? If so, how can best we help them get greater visibility?

As to what families such platforms could prioritize for first help, consider the following disturbing fact from a 2013 Lancet series on Maternal and Child Nutrition: Roughly 3 million children (5 years and younger) die a year, one every ten seconds, 8,000 a day. These deaths are either a result of lack of adequate food for the mother or child, or diseases induced by undernutrition. Half of these child deaths occur in one planetary location of greatest need: Sub-Saharan Africa. Globally, undernutrition accounts for 45% of child deaths. Over 165 million living children today are both stunted and immunocompromised because of lack of adequate nutrition in their childhood. Eradicating extreme hunger, reducing child mortality, and improving maternal health are #1, #4, and #5 of the eight UN Millennium Development Goals.
How soon will it be before some social entrepreneur or human development NGO develops an Internet of Families sensor package and Private Social Network for adopting an at-risk family and improving their lives, beginning with things like better nutrition, housing, sanitation, and security? Would you adopt an at-risk family in Sub-Saharan Africa or some other place of great need if you could participate in their lives even more easily than if they were your physical neighbors? I certainly would. Let’s get it done!
We need a platform that NGOs and families everywhere can use to nominate and monitor the most at-risk moms, dads, and kids globally. If we can stick chips in our fancy household appliances, we can network and sensor-equip our adopted at-risk families in Mali, Chad, Pakistan, and everywhere else on this beautiful and sometimes dangerous planet. I would love to be able to see webcams of what my adopted family is doing right now and has been doing this week, with video, audio, images and data all auto-summarized on a Private Social Network. So would many others who want to give aid and develop lifetime personal relationships where they can matter most, in at-risk children and their parents, with minimal interference by NGOs between us and the people in need.
I’d love my baby to grow up talking and live streaming with her adopted sisters and brothers in another country. Surely, there are legal issues to work out. And security and political issues. Perhaps these social concerns, and specifically, an unwillingness to shoulder the legal risks that would ensure, are why even our leading global aid platforms, like Kiva.org, give us no way to directly contact or support the families to whom we give our microloans. There’s one word for this level of service, which is worth repeating: lame, lame, lame! Don’t tell us the problem can’t be solved. Even if there are political blocks to offering such a personalized service in some country, it could be offered in others, and the blockers would watch their kickbacks disappear as relief flows moved to where the light and personalization is best. And over the long term, autocratic leaders can’t stop cellphones and sensors and networks from penetrating illiberal countries. The network just keeps routing around the damage, as they say.
It’s time for a better solution to the terrible pain and misery that the world’s most at-risk families face every day. You can be sure some global entrepreneurs could create both formal and informal at-risk family adoption processes that scale with exponential speed, and put our formal adoption agencies to shame.
Here’s a Vision

Imagine that 20 million of the most at-risk families around the world are identified by trusted high-reputation individuals and NGOs and paired directly with one of the 440 million households of the thirty-four OECD member countries, on an Internet of Families and Private Social Network platform that nudges them into regular intimate and rich media communication with and financial, educational, emotional, and other support of their adopted family. That’s an NGO mission, and a call to shared responsibility and transparency, that would be a worthy channel for some of our global humanitarian impulses.
Imagine next that, over ten years of scaling, one out of twenty of these 440 million households (5% of the world’s privileged) decides to sponsor an at-risk family via this Internet of Families, either from another country or from among the homeless, disadvantaged, and poor of their own country, or county. That Early Adopter group would cover 22 million of the most needy families in the world. It would probably be the most satisfying philanthropy those sponsor families ever did in their lives, and it would also promote global interdependence in ways we can’t really imagine today.
A good Global Family Adoption platform will be evidence-based in its recommendations of support, maximizing quality-adjusted life years for the at-risk family by getting us most privileged folks to focus our donations and interactions on help with maternal and child nutrition, maternal, paternal and child education, reproductive choice counseling, and a host of other issues. Video snips of our adopted family and all their sensor data will be uploaded daily to the web, to record any violence or corruption or misfortune happening to them. Then we’ll start figuring out how to fight and stop that violence and corruption via mass action. When problems happen to your family, it becomes personal. We stop bitching about it and get out there and fix the problem.
Did you know that from 2011 to 2014 Malaysia gave out 1.5 million netbooks to their children and established thousands of village wi-fi hubs in their National Broadband Initiative? More of that kind of boldness is happening around the world every year now. There’s no reason some visionary entrepreneurs and funders can’t roll out a first generation Global Family Adoption platform with an Internet of Families and a rich Private Social Network today. Millions of us privileged families are ready to donate money to establish this kind of regular intimacy with those who need it most. There’s no reason we can’t use our technology now in this incredibly empowering way.
Edits? Additions? Changes? Comments always appreciated, thanks for reading.












 Until
Until 









